Текущая ситуация и перспективы развития вакцин против болезни Ньюкасла
Болезнь Ньюкасла (БН) — одно из самых экономически разрушительных инфекционных заболеваний, поражающих птицеводческую отрасль. Вирулентный вирус болезни Ньюкасла (ВБН) может вызывать высокую смертность и серьёзные поражения тканей дыхательной, желудочно-кишечной, нервной, репродуктивной и иммунной систем домашней птицы. Благодаря строгим мерам биобезопасности и широкому применению вакцин был достигнут значительный прогресс в предотвращении заболеваемости и смертности, вызванных БН. За последние десятилетия непрерывная эволюция ВБН привела к появлению в общей сложности двадцати генотипов, и генетическая изменчивость может быть связана со вспышками заболеваний у вакцинированных кур. В некоторых странах применение новых вакцин, подобранных по генотипу, успешно подавляет циркуляцию вирулентных штаммов вируса диареи птиц в полевых условиях. Однако вирулентный вирус диареи птиц по-прежнему распространен во многих регионах мира, особенно в странах с низким и средним уровнем дохода, что негативно сказывается на средствах к существованию миллионов людей, для которых мясо птицы является основным продуктом питания. В странах, эндемичных по лихорадке денге, несмотря на вакцинацию для борьбы с заболеванием, эффективность вакцинации снижается из-за отсутствия вакцин, соответствующих генотипу вируса, которые могли бы снизить риск заражения и передачи вируса, а также из-за ненадлежащего применения вакцин на местах. Изучение характеристик существующих вакцин против лихорадки денге имеет основополагающее значение для разработки надлежащих режимов вакцинации и создания вакцин нового поколения. Поэтому в этой статье мы представляем широкий обзор коммерческих и экспериментальных вакцин против лихорадки денге, а также перспективных новых платформ для разработки вакцин нового поколения.
Болезни Ньюкасла
Болезнь Ньюкасла (БН) — это серьёзное инфекционное заболевание домашней птицы, которое существует уже почти сто лет и стало причиной как минимум четырёх панзоотий по всему миру. БН вызывается вирусами болезни Ньюкасла (ВБН). Первая панзоотия, охватившая период с 1930-х по 1960-е годы, была вызвана вирусами генотипов I, II, III и IV. Вторая панзоотия, охватившая период с конца 1960-х по 1973 год, была вызвана в основном вирусами генотипов V и VI. В 1975 году у голубей началась третья панзоотия, которая распространилась на различные регионы мира. Причиной этой панзоотии стал вирус ньюкаслской болезни генотипа VI. С конца 1980-х годов вирус ньюкаслской болезни генотипа VII появился на Дальнем Востоке и распространился по всему миру, вызвав четвёртую панзоотию. В настоящее время генотип VII NDV является эндемичным для многих стран Азии и Африки и представляет большую угрозу для птицеводческой отрасли.
NDV относится к роду Orthoavulavirus подсемейства Avulavirinae. NDV — это оболочечный вирус с одноцепочечным несегментированным геномом РНК с негативной полярностью (рис. 1A). Геном NDV кодирует шесть вирусных белков: нуклеопротеин (NP), фосфопротеин (P), матриксный (M), слияния (F), гемагглютинин-нейраминидазу (HN) и большой (L) белки (рис. 1B). На основании патогенности, наблюдаемой у восприимчивых кур, вирус ньюкаслской болезни можно разделить на три патотипа: велогенный, мезогенный и лентогенный. Штаммы с индексом внутримозговой патогенности (ICPI) ≥ 0,7 считаются вирулентными в соответствии со стандартами Всемирной организации по охране здоровья животных (МЭБ). Вирулентные штаммы вызывают системное заражение и тяжелые поражения у кур, а аминокислота в месте расщепления F-белка является основным фактором, определяющим вирулентность.
Рисунок 1
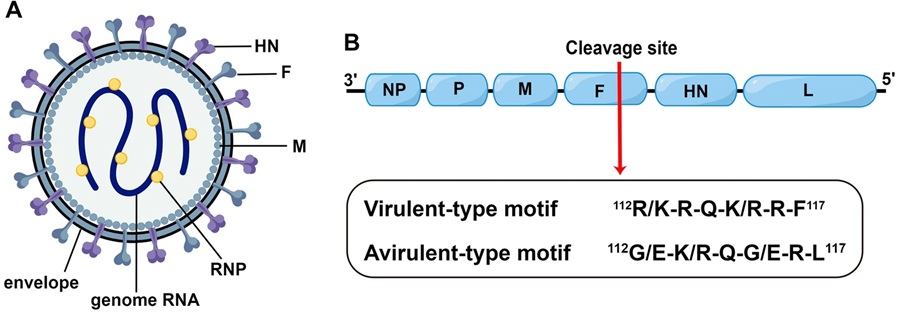
Схематическое изображение частиц и структуры генома вируса денге. Изображение структуры частиц вируса денге. HN — гемагглютинин-нейраминидаза; F — белок слияния; M — матриксный белок; RNP — рибонуклеотид-белковый комплекс. B Схематическое изображение структуры генома вируса денге. Красной стрелкой обозначен сайт расщепления белка F. Показаны репрезентативные аминокислотные мотивы сайтов расщепления вирулентного и авирулентного типов.
Живые вакцины
К живым вакцинам против ньюкаслской болезни относятся лентогенные и мезогенные вакцины. В мире наиболее широко используются лентогенные живые вакцины, а La Sota и B1 являются их репрезентативными штаммами. Эти вирусы относятся к генотипу II и имеют высокое генетическое и антигенное сходство. Лентогенные вакцины могут вызывать выработку защитных антител, хотя они различаются по тканевому тропизму и характеру репликации у кур. Штамм La Sota обладает высоким тропизмом к дыхательной системе и активно реплицируется у кур. Титры антител, вырабатываемых вакциной La Sota, как правило, высоки, поэтому эта вакцина подходит для использования в странах, где эндемичен вирулентный вирус диареи Ньюкасла. Вакцина VG/GA характеризуется двойным тропизмом к дыхательным путям и кишечнику, при этом тропизм к кишечнику выше, и она может вызывать сильный иммунитет к слизистым оболочкам. Кроме того, вакцина B1 обладает очень низкой вирулентностью и безопасна для цыплят, поэтому её обычно используют при низком уровне заражения или у цыплят. Другой тип лентогенных живых вакцин основан на генотипе I, и V4 и I-2 являются представителями этого типа. Эти штаммы обладают низкой вирулентностью и безопасны для кур всех возрастов. Кроме того, V4 и I-2 — это типичные термостабильные вакцины, обладающие уникальным преимуществом: их можно использовать в отдалённых районах страны с ограниченным доступом к холодовой цепи, а также вводить с питьевой водой и пищей.
Живые вакцины против ньюкаслской болезни безопасны, поскольку вакцинные штаммы обладают низкой вирулентностью или не обладают ею вовсе и обычно вызывают у кур лишь незначительные поствакцинальные реакции. Кроме того, живые вакцины могут вызывать слизистый, гуморальный и клеточный иммунитет и могут вводиться путём распыления или с питьевой водой. Однако при определённых условиях живые вакцины могут вызывать нежелательные поствакцинальные реакции, такие как замедление роста, лёгкие респираторные симптомы и даже смертность, а также повышать восприимчивость к другим патогенам. Для транспортировки и хранения этих вакцин требуется холодильное оборудование. Кроме того, живые вакцины обычно вводят молодняку, и наличие материнских антител (МА) может снижать их эффективность. Таким образом, для обеспечения эффективности вакцины необходимо вводить живые вакцины против ньюкаслской болезни в нужный момент, определяемый по уровню МА.
Другой тип живых вакцин основан на мезогенных штаммах, таких как Mukteswar (генотип III), Komarov (генотип II) и Roakin (генотип II). Эти штаммы обладают высокой вирулентностью и могут вызывать гибель молодняка. Мезогенные живые вакцины обладают высокой иммуногенностью и могут вызывать быстрый и продолжительный иммунный ответ. Эти вакцины можно использовать для ревакцинации или экстренной вакцинации взрослых кур. Однако, поскольку МЭБ определяет мезогенные вирусы как вирулентные, а также сообщалось о повышенной вирулентности этого типа вакцин, мезогенные живые вакцины во многих странах в значительной степени запрещены.
Живые противораковые вакцины производятся из куриных яиц, не содержащих специфических патогенов (SPF), с эмбрионированными яйцами (ECES). Основное семя вакцины инокулируют в аллантоисную полость 9-11-дневных ECE, а аллантоисные жидкости собирают после инкубации. После прохождения серии тестов по контролю качества аллантоиновые жидкости, дополненные стабилизаторами, подвергаются лиофилизации. Этот процесс крайне важен для сохранения содержания антигена и срока годности вакцин. Кроме того, определение 50-процентной эмбриональной инфекционной дозы (ЭИД50) является ключевым этапом для оценки уровня антигена и эффективности живых вакцин. Исследования подтвердили взаимосвязь между дозой вакцины (в ЭИД50) и эффективностью защиты. В экспериментальных условиях цыплята SPF, вакцинированные живой вакциной La Sota в дозе 104–105 ЭИД50, были защищены от заболеваемости и смертности, хотя заражение и репликация вируса, вызвавшего заболевание, не были снижены. Для обеспечения полной защиты и снижения выделения вируса живыми вакцинами требуется доза в 106 ЭИД50. Таким образом, утверждается, что трёх целей вакцинации против ньюкаслской болезни можно достичь при достаточно высокой дозе вакцины. Однако на практике увеличение дозы вакцины приводит к росту затрат на борьбу с заболеванием, что делает её использование в птицеводстве нецелесообразным.
Основная проблема, связанная с современными живыми вакцинами, заключается в том, что они не соответствуют доминирующим вирусам. Несмотря на то, что вирус диареи крупного рогатого скота относится к одному серотипу, между обычными живыми вакцинами и вирусами, присутствующими в природе, наблюдаются значительные генетические и антигенные различия. Многие исследования показали, что гомология аминокислотных последовательностей белков F и HN между штаммом La Sota и штаммами генотипа VII составляет 87–89 % и 87–88 % соответственно. Аналогичным образом, в Латинской Америке наиболее распространённые генотипы, поражающие птицеводческие хозяйства, соответствуют генотипам V, VI, VII, XII и XVI, а идентичность между La Sota и преобладающими штаммами составляет 87–89 %. По этому вопросу существуют две противоположные точки зрения. В некоторых отчётах отмечается, что антигенная разница между вакцинами и преобладающими штаммами не является основной причиной вспышек заболеваний в полевых условиях. Вместо этого причиной низкой эффективности живых вакцин может быть слабый иммунитет у животных, вызванный недостаточной практикой вакцинации. Эти результаты подчёркивают важность достаточной вакцинации для обеспечения эффективности живых вакцин в полевых условиях. С другой стороны, введение живых вакцин против НЯ, гомологичных циркулирующим вирусам, способствует снижению выделения вируса в стаде. Одна экспериментальная и две коммерческие живые рекомбинантные вакцины против генотипа VII обеспечивали лучшую защиту от полевых изолятов у кур, чем La Sota. Соответственно, живая рекомбинантная вакцина против НЯ, экспрессирующая гены F и HN из генотипа V, была более эффективной в снижении выделения вируса, чем штамм La Sota. Кроме того, убедительные доказательства, подтверждающие этот вывод, были получены в ходе исследований инактивированных вакцин против ящура, о чём подробно говорится ниже. С точки зрения механизма, недавно была предложена модель, согласно которой вирус вакцины может быть нейтрализован низкими титрами антител, выработанных гомологичной вакциной, в то время как для нейтрализации полевых штаммов, содержащих вариации в основных нейтрализующих эпитопах, требуются более высокие титры антител. Эта модель подтверждает необходимость разработки вакцин, гомологичных полевым вирусам. Таким образом, для обеспечения надёжной защиты в полевых условиях необходимы как вакцины, соответствующие генотипу, так и надлежащие процедуры вакцинации.
Основная проблема при создании живых вакцин против вирулентных штаммов вируса диареи крупного рогатого скота, соответствующих генотипу, заключается в том, как обеспечить их антигенность и безопасность. Для создания живых вакцин, соответствующих генотипу, используются две основные стратегии. Некоторые вирулентные штаммы были ослаблены с помощью обратной генетики и модифицированы для получения живых вакцин. Вирулентные штаммы ослабляются путем модификации сайта расщепления F, а дополнительные специфические мутации в белке L могут дополнительно гарантировать безопасность ослабленного вируса. Ни одна живая аттенуированная вакцина, созданная с использованием этой стратегии, пока не поступила в продажу. Другая стратегия заключается в замене защитных антигенов F и HN лентогенного вируса соответствующими белками из распространённых вирулентных штаммов, при этом сайт расщепления F обычно мутируется. Несколько живых рекомбинантных вакцин против энтеровируса человека этого типа поступили в продажу в Корее (живая вакцина Himmvac Dalguban N (Plus)), Египте (живая аттенуированная вакцина RINNOVAC™ELI-7) и Мексике (Genovax N5). Кроме того, на эффективность рекомбинантных вакцин может влиять происхождение лентогенных основ NDV. Следует отметить, что безопасность этих живых вакцин на основе вирулентных вирусов должна тщательно контролироваться, даже если исследования в лабораторных условиях с чётким контролем подтвердили их безопасность для подопытных животных.
Инактивированные вакцины
Инактивированные вакцины также широко используются для борьбы с болезнями. Аллантоидные жидкости получают из эмбрионов, заражённых исходным семенем, и инактивируют формалином или β-пропиолактоном. Для полной потери инфекционности аллантоидные жидкости необходимо инактивировать несколько раз. Для приготовления эмульгированных вакцин в инактивированные аллантоидные жидкости добавляют адъюванты, например минеральное масло. Стоимость производства инактивированных вакцин относительно высока. В качестве исходного материала обычно используются лентогенные штаммы вируса ньюкасла, в том числе La Sota, Ulster и B1, из-за их высокой вирусной активности в эмбрионах кур-несушек. Инактивированные вакцины безопасны, поскольку вирусы не могут размножаться и распространяться среди вакцинированных кур. Инактивированные вакцины вводятся индивидуально парентеральным способом, например внутримышечно или подкожно (п/к), что делает процесс трудоёмким и длительным. Эти вакцины в основном вызывают высокий и продолжительный гуморальный иммунитет, но плохо стимулируют клеточный иммунитет или иммунный ответ слизистых оболочек.
Как и в случае с живыми вакцинами, эффективность инактивированных вакцин зависит от степени их соответствия циркулирующим вирусам. В большинстве стран, где болезнь Ньюкасла является эндемическим заболеванием, птицам вводят инактивированные вакцины на основе штамма La Sota. Примечательно, что аминокислотная идентичность белков F и HN между традиционными вакцинными штаммами и доминирующими вирусами болезни Ньюкасла составляет 87–89 % и 87–88 % соответственно. Инактивированные вакцины на основе La Sota вызывают значительно более низкие титры HI-антител против гетерологичного вируса, чем против гомологичного антигена. Более того, куры, вакцинированные обычными вакцинами, полностью защищены от заболеваемости и смертности, хотя они всё равно могут выделять большое количество вируса. Эти результаты могут объяснить частое возникновение атипичной формы НЯ в стадах, вакцинированных традиционными вакцинами.
В отличие от живых аттенуированных вакцин, проблему соответствия генотипу в инактивированных вакцинах против НЯ легче решить с помощью обратной генетики. Рекомбинантные вакцины, специфичные для генотипа VII вируса НЯ, были созданы с помощью различных стратегий. Некоторые из этих вакцин-кандидатов уже поступили в продажу в разных странах. Корейская команда разработала рекомбинантную вакцину-кандидат на основе генотипа VII (KBNP-C4152R2L), экспрессируя гены F и HN в основе La Sota, и эта вакцина-кандидат поступила в продажу в виде живой и инактивированной вакцин. Исследования подтвердили, что эта вакцина обеспечивает более надёжную защиту, чем La Sota, с точки зрения снижения выделения вируса Кроме того, наша команда разработала аттенуированную вакцину против диареи крупного рогатого скота генотипа VII (A-VII) путем мутации аминокислотного мотива в сайте расщепления F. По сравнению с вакциной La Sota, инактивированная вакцина A-VII вызывает более быструю и сильную реакцию антител, полностью защищая цыплят от заражения вирусом генотипа VII и значительно снижая выделение вируса. Примечательно, что снижение выделения вируса впервые было включено в Китайскую ветеринарную фармакопею в качестве ключевого стандарта для проверки эффективности вакцин против ньюкаслской болезни. На 5-й день после заражения не менее чем у 7 из 10 вакцинированных цыплят в мазках из клоаки не должно быть обнаружено вируса [58]. Вакцину A-VII также можно использовать для борьбы с болезнями у гусей. После коммерциализации A-VII в 2014 году и его широкого применения в птицеводстве заболеваемость НЯ резко снизилась, а циркуляция вирулентного генотипа VII вируса диареи крупного рогатого скота в птицеводческих хозяйствах Китая практически прекратилась (согласно данным Китайского официального ветеринарного бюллетеня). Вакцины, соответствующие генотипу, гомологичны преобладающим вирусам и более эффективны в предотвращении выделения вируса. Таким образом, возникает вопрос о том, какие генетические или антигенные вариации чаще всего возникают у полевых вирусов под воздействием гомологичных вакцин. Недавнее исследование, проведённое с использованием анализа нейтрализации сыворотки in vitro, показало, что не существует существенных различий в вариативности генов F и HN генотипа VII NDV под давлением отбора гомологичных и гетерологичных вакцин. Это означает, что применение вакцин, соответствующих генотипу, может не повлиять на вариативность генотипа VII NDV.
Помимо вакцин для кур и гусей, существует большой спрос на вакцины против ньюкаслской болезни, предназначенные для голубей. В отличие от вирусов ньюкаслской болезни, имеющих другое происхождение, вирусы, поражающие голубей, также известные как парамиксовирус голубей (PPMV)-1 (в основном генотип VIb), специфичны для голубей. Этот генотип также является поздним генотипом, демонстрирующим высокую генетическую и антигенную изменчивость по сравнению с традиционными вакцинными штаммами [60]. Инактивированные вакцины на основе штамма La Sota или Ulster могут обеспечить полную защиту от инфекции PPMV-1, тогда как они индуцируют более низкие титры HI против PPMV-1 и не снижают выделение вируса у голубей. Для голубей доступны только инактивированные вакцины, такие как AVIPRO 111 PMV1 и Nobilis Paramyxo P201 (данные the PoultryMed). Кроме того, с помощью обратной генетики можно модифицировать PPMV-1, чтобы повысить его безопасность, антигенность и выход вируса, что может стать перспективным направлением в разработке вакцин против орнитозной диареи голубей.
Вакцины на основе вируса оспы кур
Создание поливалентных вакцин может расширить спектр вакцин и повысить эффективность вакцинации в птицеводстве. Традиционно для производства поливалентных вакцин для птицы требуется определённая концентрация антигенов для поддержания достаточной дозы каждого антигена, что, несомненно, усложняет весь процесс и увеличивает его стоимость. В качестве альтернативы перспективными системами для разработки бивалентных или поливалентных вакцин для птицы стали платформы на основе вирусных векторов. Гены защитных антигенов патогенов-мишеней встраиваются в геном вируса-переносчика с помощью молекулярно-биологических технологий, и рекомбинантные вирусы экспрессируют чужеродные антигены в процессе репликации. По сравнению с традиционными поливалентными вакцинами, вакцины на основе вирусных векторов имеют ряд преимуществ, в том числе возможность совместной экспрессии нескольких защитных антигенов в одном вирусном векторе, отсутствие необходимости в концентрации антигенов, индукция гуморального, клеточного или мукозального иммунитета, высокая генетическая стабильность и безопасность.
С 1990-х годов учёные встраивали гены F или HN вируса ньюкаслской болезни в геном вируса оспы кур (ВОК), и полученные в результате рекомбинантные вакцины могли защищать кур от заражения вирулентными штаммами вируса ньюкаслской болезни. На рынке доступны как минимум две коммерческие вакцины против вируса ньюкаслской болезни на основе ВОК, в том числе TROVAC®-NDV от Boehringer Ingelheim и VECTORMUNE® FP-N от Ceva. Вирус птичьего гриппа — самый крупный вирус животного происхождения, и его геном обладает высокой способностью к включению чужеродных генов. Кроме того, вирус птичьего гриппа имеет ограниченный круг хозяев и поэтому безопасен для других видов животных. Вирус птичьего гриппа обычно размножают в эмбрионах кур, полученных от здоровых несушек, или в фибробластах куриных эмбрионов, а затем собирают хориоаллантоисную мембрану или клеточные культуры для лиофилизации с добавлением стабилизатора. Иммунизация вакцинами против НЯ на основе вируса птичьего гриппа проводится путем подкожной инъекции или прокола перепонки крыла, поэтому их массовое применение невозможно. Кроме того, наличие высоких титров MDA против вируса инфекционного бронхита кур у промышленных кур значительно снижает эффективность вакцин на основе вируса инфекционного бронхита. Эти недостатки вируса инфекционного бронхита означают, что его использование для доставки вакцин против ньюкаслской болезни сильно ограничено.
Вакцины на основе вируса герпеса индеек
Вакцины против ньюкаслской болезни на основе вируса герпеса индеек (ВГВИ) являются успешным примером коммерческих вирусных вакцин для домашней птицы. ВГВИ, также известный как вирус болезни Марека 3-го серотипа (ВБМ), является одним из наиболее широко используемых вирусов для создания новых векторных вакцин. В начале 1990-х годов ген F вируса ньюкаслской болезни был встроен в геном ВГВИ, и полученный рекомбинантный вирус может обеспечивать двойную защиту от вируса ньюкаслской болезни и вируса болезни Марека. На рынке представлены четыре коммерческие вакцины против НЯ на основе HVT-вектора, в том числе Vectormune® ND от Ceva, Innovax-ND от MSD, Poulvac Procerta HVT-ND от Zoetis и NEWXXITEK™ HVT + ND от Boehringer Ingelheim (данные PoultryMed). Вакцина Vectormune® ND вызывает замедленный, но стойкий иммунный ответ и обеспечивает хорошую защиту от вируса НЯ. Ген F штамма I NDV D26-76 экспрессируется в штамме HVT FC126. Несмотря на то, что вирус-донор гена F отличается от циркулирующих вирусов, вакцина Vectormune® ND может обеспечить защиту от гетерологичных генотипов (IV, V и VII) и снизить выделение вируса . В частности, вакцина против НЯ на основе HVT-вектора более эффективно снижает выделение вируса при внутримышечном введении генотипа VII вируса НЯ по сравнению с интраназальным введением, что позволяет предположить, что вакцина вызывает сильный системный иммунитет. В регионах с низким риском НЯ введение вакцины Vectormune® против НЯ путем интродукции 18-дневных эмбрионов или подкожного введения суточным цыплятам может обеспечить достаточную защиту. В регионах со средним и высоким риском заражения оспой обезьян рекомендуется проводить первичную иммунизацию с использованием вакцины Vectormune® против оспы обезьян, а через 2–3 недели — ревакцинацию живыми вакцинами для обеспечения надежной защиты. К преимуществам вектора HVT относятся следующие:
(1) геном HVT большой и может вмещать множество чужеродных генов;
(2) Влияние MDA на HVT низкое, что позволяет проводить раннюю вакцинацию цыплят;
(3) массовое введение вакцин с HVT-вектором может быть выполнено в инкубаториях путем инъекции in ovo или внутривенно суточным цыплятам;
(4) Вакцины с HVT-вектором могут обеспечивать защиту на протяжении всей жизни из-за персистирующей инфекции HVT;
(5) Вакцины с HVT-вектором могут индуцировать как антителный, так и клеточный иммунитет. Эти сильные стороны лежат в основе успеха вакцин с HVT-вектором во всем мире.
Кроме того, инструмент для редактирования генома CRISPR/Cas9 можно использовать в качестве простого и быстрого способа разработки рекомбинантных вакцин на основе вируса птичьего гриппа. Однако у вируса птичьего гриппа есть некоторые ограничения, такие как замедленное формирование иммунитета и невозможность введения второй вакцины на основе вируса птичьего гриппа в том же стаде. Кроме того, поскольку вакцины против гемофильной инфекции выпускаются в виде лиофилизированных препаратов или замороженных клеточных суспензий, для их хранения требуется холодильная цепь или жидкий азот, что может ограничивать их применение в регионах, где нет необходимых условий.
Векторные вакцины на основе NDV
NDV является хорошим вектором для создания бивалентных или мультивалентных вакцин против НЯ и других инфекционных заболеваний домашней птицы. У вируса денге есть ряд преимуществ при использовании в качестве вектора для вакцины:
(1) геном вируса денге составляет ~15 килобаз, что упрощает молекулярные манипуляции;
(2) вирус денге обладает высокой вирусной продуктивностью в клеточных культурах;
(3) вирус денге может стабильно встраивать и экспрессировать чужеродный ген;
(4) вирус денге обладает низким риском обмена генами и рекомбинации;
(5) вирус денге может вызывать мукозальный, гуморальный и клеточный иммунитет;
(6) вакцины на основе вируса денге можно вводить методом массовой вакцинации.
Были созданы многочисленные вакцины-кандидаты на основе векторов NDV, включая кандидаты, нацеленные на вирус птичьего гриппа (AIV), IBDV и вирус инфекционного бронхита (IBV). Две вакцины против H5 подтипа AIV, основанные на переносчиках NDV, были коммерциализированы в Китае и Мексике.
Большинство бивалентных вакцин основаны на лентогенных вирусах денге, а некодирующая область между генами P и M является оптимальным местом для встраивания чужеродных генов. Общая стратегия создания вакцин заключается во встраивании чужеродных генов в качестве независимой единицы транскрипции в геном вируса денге, а транскрипционные сигналы, включая концевые, межгенные и стартовые последовательности генов, добавляются перед чужеродными генами. Рекомбинантный вирус денге можно восстановить с помощью обратной генетики, а затем систематически оценивать репликацию вируса, экспрессию чужеродных белков, вирулентность, генетическую стабильность и эффективность на животных.
Несмотря на перспективность вакцин на основе вируса ньюкасла, серьёзную озабоченность вызывает интерференция со стороны вируса инфекционного бронхита кур. При использовании общей стратегии создания вакцин чужеродные антигены встраиваются в оболочку вирусных частиц. Таким образом, антитела против вируса ньюкасла, а также чужеродные антигены могут влиять на действие векторных вакцин. Подтверждающие данные были получены в ходе предыдущего исследования, продемонстрировавшего более сильное влияние антител против H5 на вакцины против H5 на основе вируса ньюкасла, чем влияние антител против вектора у кур. Исследователи приложили немало усилий, чтобы решить проблему интерференции MDA, и разработали две основные стратегии: «маскировка антигена» и «антигенный обман». Во-первых, что касается интерференции с вектором, была разработана стратегия «маскировки антигена», основанная на низкой перекрестной реактивности между вирусом диареи крупного рогатого скота и другими серотипами парамиксовируса птиц (APMV). Гены F и HN вируса ньюкаслской болезни заменяются соответствующими генами вируса инфекционного бронхита кур-2 или -8, и на полученный химерный вектор не влияют уже имеющиеся у кур антитела к вирусу ньюкаслской болезни. Рекомбинантные вакцины на основе химерных векторов обладают высокой иммуногенностью и эффективностью у кур с антителами к вирусу инфекционного бронхита кур. Во-вторых, недавно была предложена стратегия «антигенной приманки» для борьбы с интерференцией чужеродных антигенов. На примере подтипа H5 AIV было показано, что гемагглютинин (HA) экспрессируется в полноразмерной и секретируемой версиях в векторе NDV. Секретируемая форма действует как приманка, которая может поглощать часть уже существующих антител, специфичных к HA. Таким образом, заякоренный в мембране белок гемагглютинина защищён от связывания антителами к гемагглютинину, что позволяет этим антигенам вызывать специфический иммунитет. Поэтому недавно мы предположили, что сочетание стратегий «маскировки антигена» и «приманки для антигена» может стать эффективным способом разработки новых векторных вакцин против NDV, устойчивых к MDA.
Рекомбинантные субъединичные вакцины
В настоящее время почти все вакцины против ньюкаслской болезни производятся в клеточных батареях, которые представляют собой традиционную, отработанную систему производства вакцин для домашней птицы. В клеточных батареях легко осуществлять крупномасштабное культивирование, а вирус ньюкаслской болезни имеет высокий выход в клеточных батареях. Однако у системы на основе яиц есть и недостатки, в том числе высокая стоимость, нестабильные поставки (особенно во время вспышек заболеваний), образование большого количества биологических отходов, высокое энергопотребление и выбросы углекислого газа при переработке биологических отходов. Поэтому необходимо разрабатывать альтернативные платформы для производства вакцин для домашней птицы.
Система экспрессии бакуловируса (BEVS) является многообещающей альтернативой системе на основе куриных яиц. С помощью BEVS производится несколько коммерческих вакцин для людей и животных. Рекомбинантный бакуловирус, экспрессирующий чужеродные антигены, генерируется и размножается в клетках насекомых для крупномасштабного производства. У BEVS есть несколько существенных преимуществ:
(1) функция посттрансляционной модификации в клетках насекомых подходит для экспрессии вирусных гликопротеинов;
(2) крупномасштабное культивирование клеток насекомых в бессывороточной среде позволяет получать большое количество антигенов с минимальными затратами;
(3) бакуловирус обладает высокой специфичностью к хозяину и безопасен для позвоночных животных;
(4) геном бакуловируса обладает высокой способностью к совместной экспрессии нескольких чужеродных антигенов;
(5) бакуловирус обладает адъювантным эффектом.
На рынке нет вакцин против НЯ, основанных на BEVS. На доклиническом этапе белки F или HN экспрессируются в клетках sf9, а антигены собираются для приготовления субъединичных вакцин. Благодаря наличию основных защитных антигенов иммуногенность и эффективность субъединичных вакцин против НЯ сопоставимы с эффективностью цельновирусных инактивированных вакцин. Следует отметить, что процесс дополнительной очистки антигена и связанное с ним увеличение затрат вызывают опасения в отношении субъединичных вакцин для домашней птицы. Недавнее исследование показало, что очистка антигена для субъединичных вакцин может быть необязательной. Субъединичная вакцина против птичьего гриппа подтипа H7N9, полученная в клетках насекомых в виде неочищенного антигена, обладает высокой иммуногенностью и эффективностью у кур. Учитывая необходимость контроля затрат, производство субъединичных вакцин на основе неочищенных антигенов, экспрессируемых в клетках насекомых, может быть приемлемым для вакцин против птичьего гриппа.
Системы экспрессии на основе растений представляют собой привлекательную платформу для производства субъединичных вакцин для домашней птицы. Защитные антигены F и/или HN экспрессируются в различных растениях, таких как N. benthamiana, Zea mays, Solanum tuberosum и Oryza sativa. Эти антигены могут вызывать выработку специфических антител или клеточный иммунитет у кур или мышей. Когда Шахид и др. выразили белок HN в кукурузе и скормили курам листья и семена кукурузы, у них выработался специфический для NDV иммунный ответ в виде антител. Было показано, что рекомбинантный белок F, полученный из риса, вызывает выработку антител и обеспечивает полную защиту от инфекции, вызванной вирусом NDV генотипа VII. Более того, вакцина на основе субъединицы F позволяет дифференцировать инфицированных и вакцинированных животных (DIVA) путем выявления антител, специфичных для HN. Таким образом, системы на основе растений являются перспективными для создания новых вакцин против диареи из-за таких их преимуществ, как низкая стоимость, отсутствие или минимальная потребность в очистке, экологичность и возможность перорального введения.
Вакцины на основе вирусоподобных частиц
Другим типом экспериментальной вакцины против ящура является вакцина на основе вирусоподобных частиц (ВПЧ). ВПЧ по своей морфологии и размеру напоминают естественные вирусные частицы, но не содержат нуклеиновых кислот. Вакцины на основе ВПЧ безопасны для животных-мишеней, поскольку не могут размножаться в организме хозяина. В ВПЧ включены защитные антигены, которые могут стимулировать клеточный иммунитет и выработку антител. Белок М является основной движущей силой сборки и почкования вируса ящура. Экспрессия одного М-белка достаточна для сборки VLP, и, таким образом, М-белок является обязательным для производства VLP NDV. Белки F, HN и M экспрессируются в BEV для сборки VLP, и VLP очищаются из образцов для приготовления вакцин. Сюй и др. создали вакцину-кандидат VLP против NDV генотипа VII путем коэкспрессии генов M, F и HN в клетках sf9. Было показано, что VLP-вакцина с адъювантом к квасцам вызывает более длительный период защиты и более короткий период выделения вируса, чем цельновирусная инактивированная вакцина La Sota при заражении штаммом NDV генотипа VII. Кроме того, VLP NDV также могут быть использованы в качестве платформы для создания вакцин против патогенов человека и сельского хозяйства. Поскольку сборка вирусоподобных частиц зависит от взаимодействия различных вирусных белков, необходимо оптимизировать экспериментальные параметры, чтобы получить максимальное количество вирусоподобных частиц. Для приготовления вакцин на основе вирусоподобных частиц требуется очистка антигенов.
Вакцины для дифференциации инфицированных и вакцинированных животных
Конечной целью борьбы с инфекционными заболеваниями животных является искоренение болезней. Для достижения этой цели крайне важна стратегия DIVA. В странах, где ящур является эндемическим заболеванием, вакцины DIVA помогают отслеживать циркуляцию вируса в вакцинированных стадах. В странах, где ящур хорошо контролируется с помощью программ вакцинации, например в Китае, вакцины DIVA имеют большое значение для дальнейшего искоренения болезни.
Поскольку все современные вакцины против НД содержат цельновирусные компоненты, антитела, индуцируемые этими вакцинами, нелегко отличить от антител, индуцируемых вирусной инфекцией, с помощью традиционных серологических тестов. Основываясь на вышеупомянутых профилях различных вакцин против ND, вакцины, управляемые вирусом, субъединичные вакцины и вакцины VLP, могут использоваться в качестве вакцин DIVA, поскольку они содержат только белки F и / или HN (тогда как VLP также содержат белок M.). Следовательно, DIVA может быть проведена путем выявления антител против внутренних вирусных белков, таких как белок NP, P или L. Например, как описано выше, субъединичная вакцина на основе белка F, полученная из риса, обладает высокой защитной способностью, а DIVA можно провести, обнаружив HN-специфические антитела с помощью иммунохроматографических полосок в течение 10 минут. Кроме того, учитывая низкую перекрестную реактивность между NDV и другими APMV, Питерс и др. создали рекомбинантный NDV с химерным белком HN, состоящим из глобулярной головки APMV-4 и стебельчатой области NDV. Этот вирус полностью защищает от вирулентного NDV. Что еще более важно, используя APMV-4-специфичный HI-анализ и ELISA, нацеленный на эктодомен HN, была достигнута четкая дискриминация между сыворотками, индуцируемыми рекомбинантной вакциной и вакциной La Sota. В предыдущем исследовании сообщалось о стратегии удаления эпитопов для создания вакцины против NDV, несущей маркер DIVA. Иммунодоминантный эпитоп из 18 аминокислот (IDE) в белке NP был заменен В-клеточным эпитопом гликопротеина S2 вируса гепатита мыши (MHV). ИФА против IDE NP и эпитопа MHV S2 может достичь цели DIVA. Все стратегии, описанные выше, представляют потенциал для создания вакцин DIVA ND. Примечательно, что идеальные вакцины DIVA должны быть высокоэффективными и содержать доминирующие маркеры, позволяющие проводить специфическое, чувствительное и эффективное выявление.
Иммуномодуляторы для вакцин против НЯ
Цитокины являются важнейшими компонентами врождённого и адаптивного иммунного ответа и поэтому могут использоваться в качестве иммуностимулирующих адъювантов при подготовке вакцин. Многие исследования показали, что одновременная доставка куриных цитокинов, таких как интерлейкин (IL)-1β, IL-12, IL-18 и гранулоцитарно-моноцитарный колониестимулирующий фактор (ГМ-КСФ), различными способами может значительно повысить иммуногенность и эффективность вакцин против ящура. Здесь мы сосредоточимся на обсуждении стратегии экспрессии цитокинов в остове NDV как чужеродного гена. Zhang et al. сообщалось, что рекомбинантные NDV, экспрессирующие IL-2, IL-15 или GM-CSF, вызывали быстрый ответ антител у цыплят (на 8-й день после вакцинации) и обеспечивали полную защиту от вирулентной инфекции NDV. В частности, вирус, экспрессирующий GM-CSF, оказался устойчивым к воздействию MDA. Авторы предположили, что экспрессия цитокиновых адъювантов может эффективно стимулировать созревание Т-клеток для активации В-клеточных реакций. Кроме того, ослабленный вирус ньюкаслской болезни, экспрессирующий интерферон (ИФН)-γ, был протестирован в качестве живой вакцины при иммунизации in ovo и в качестве инактивированной вакцины при инъекционном введении. К сожалению, независимо от способа введения, интерферон-гамма не смог повысить иммуногенность вакцины против ньюкаслской болезни. Кроме того, вирулентная версия вируса ньюкаслской болезни, экспрессирующая интерферон-гамма, проявляла пониженную патогенность у 4-недельных цыплят, о чём свидетельствует отсутствие смертности, снижение тяжести заболевания и выделения вируса. Эти результаты позволяют предположить, что интерферон-гамма может действовать как противовирусный фактор и не усиливает иммуногенность вируса. Интересно, что два независимых исследования выявили важную роль антисмыслового гена IL-4 в регуляции иммуногенности вируса диареи крупного рогатого скота В совокупности данные этих двух исследований показали, что овариальная иммунизация ослабленными вирусами ньюкаслской болезни, несущими антисмысловой ген IL-4, вызывает раннюю реакцию антител и обеспечивает высокий уровень защиты у кур, несущих яйца с высокими титрами MDA. Что ещё более важно, эти вакцины значительно повышают выживаемость цыплят и их прирост в весе, что делает их идеальными овариальными вакцинами против ньюкаслской болезни. Поскольку антисмысловая последовательность гена IL-4 встроена в геном вируса ньюкаслской болезни, белок IL-4 не экспрессируется во время репликации вируса. Вместо этого наблюдаемые фенотипы рекомбинантного вируса НDV могут быть связаны с наличием антисмысловой РНК IL-4, которая вырабатывается вирусом в процессе репликации. Однако влияние антисмысловой РНК IL-4 на иммунный ответ хозяина и иммуногенность вируса до сих пор неясно. Недавно Лю и др. обнаружили, что экспрессия гена 12–2, стимулируемого интерфероном, приводит к ослаблению вируса и повышению иммуногенности вакцины против энцефалита, что может быть связано с регуляторной активностью этого гена в отношении врожденного и адаптивного иммунного ответа. Таким образом, экспрессия цитокинов или продуктов генов, связанных с цитокинами, в вирусе нематод в качестве молекулярных адъювантов или регуляторов может стать перспективным способом повышения иммуногенности и способности вакцин против нематод преодолевать влияние микобактерий.
Перспективы борьбы с заболеваниями в разных странах
В зависимости от конкретной ситуации с заболеваемостью и риска возникновения заболеваний страны можно разделить на три категории, и в соответствии с этим предлагаются различные стратегии вакцинации и разработки вакцин в будущем.
Уровень 1: страны, не эндемичные по НЯ. К этому уровню относится большинство развитых стран. Благодаря строгим мерам биобезопасности и политике искоренения заболевания случаи НЯ происходят редко, и риск распространения вируса в этих странах низок. Таким образом, программы плановой вакцинации эффективны для профилактики заболевания и борьбы с ним в коммерческих хозяйствах. Для дальнейшего повышения эффективности вакцинации против НЯ крайне важно разработать новые вакцины против НЯ, подходящие для иммунизации в инкубаториях, например интраокулярные вакцины для бройлеров. Кроме того, следует рассмотреть возможность разработки вакцин против НЯК на основе клеток, поскольку производство традиционных вакцин сдерживается нехваткой яиц, а вакцины для домашней птицы можно производить более экологичным способом.
Уровень 2: эндемичные по НЯ, но хорошо контролируемые страны. Типичным примером такой страны является Китай. В последние десятилетия НЯ была эндемичным заболеванием в этих странах, но в последнее время болезнь удалось взять под контроль благодаря применению вакцин, соответствующих генотипу вируса, и соблюдению мер биобезопасности. Крайне важно продолжать текущие программы вакцинации и разрабатывать новые вакцины DIVA, которые необходимы для отслеживания распространения вируса среди домашней птицы и искоренения болезни в долгосрочной перспективе. Кроме того, чтобы соответствовать темпам развития птицеводческой отрасли, необходимо пересмотреть стратегию вакцинации. Поэтому большим спросом пользуются вакцины нового поколения против ньюкаслской болезни, которые можно вводить массово на инкубаторных станциях, например с помощью инъекций в яйцо и распыления.
Уровень 3: страны, эндемичные по НЯ. Большинство развивающихся стран можно отнести к этому уровню. Из-за неэффективной политики биобезопасности, ненадлежащего хранения или применения вакцин поголовье домашней птицы подвергается высокому риску заражения вирулентными вирусами в полевых условиях. Таким образом, важно внедрять строгие меры биобезопасности, разрабатывать новые вакцины, соответствующие доминирующим вирусам, и правильно применять их в полевых условиях, поскольку эти стратегии могут снизить количество выделяемых и циркулирующих в поголовье вирусов. Кроме того, в некоторых развивающихся странах из-за нехватки холодильного оборудования и квалифицированных ветеринаров фермеры могут отдавать предпочтение термоустойчивым вакцинам против ящура, которые легко вводить.
В отличие от ситуации в некоторых развитых странах, в большинстве развивающихся стран продолжается распространение НЯ, что вызывает серьёзную обеспокоенность в контексте продовольственной безопасности, связанной с птицеводством. Широкое применение доступных в настоящее время вакцин играет важную роль в профилактике заболеваний и борьбе с ними как в развитых, так и в развивающихся странах. Поскольку вирус НЯ очень заразен, а дикие птицы иногда могут быть его носителями, не заболевая при этом, вспышки НЯ могут происходить в любом месте, где разводят домашнюю птицу. Кроме того, ситуация с заболеваемостью в разных странах и на птицефабриках отличается, что подчёркивает важность применения вакцин против НЯ в соответствии с назначением и разработки вакцин против НЯ нового поколения.

- Как отдельные сильнодействующие бактериоцины изменяют микробиом кишечника цыплят-бройлеров
- Болезни яиц
- Причины вызывающие диарею у кур. Лечение диареи у кур
- Пробиотики и ферменты в птицеводстве: отличная комбинация
- Новые подходы к диагностике ассоциированных инфекций у кур
- Морфолого-технологическая характеристика трахеи африканского страуса
- Постбиотик как эффективное средство в борьбе с сальмонеллезом птиц
- Болезни птиц: как предотвратить эпизоотию и избежать огромных убытков
- Противовирусная эффективность чесночного масла против вируса Ньюкасла
- Влияние антибиотика и пробиотика на качество мяса и субпродуктов цыплят-бройлеров
- Как сложить пазл против сальмонеллы – чему мы можем поучиться у Европы?
- Каплунирование сельскохозяйственной птицы
- Как выбрать действительно качественный гепатопротектор для птицы?
- Исследование многофункциональности вирусных гликопротеинов F и HN в жизненном цикле и патогенезе вируса болезни Ньюкасла
- Различия в применении живых и аттенуированных вакцин против кокцидиоза у птиц
- Линька у птиц, кур, выщипывание перьев
- Обсуждение достижений в борьбе с болезнью Марека на Конференции ветеринаров в США
- Ветеринарная защита в птицеводстве. Контроль заболеваний.
- Биозащита птицефермы
- Острая токсичность норфлоксацина для цыплят
- Гангренозный дерматит у кур и индеек